Via Professor Jessica Conway’s really nice paper and complete notes in her supplementary file, I’m redoing a calculation for my own model with my own notation so that I can see how it works.
First let me write down the full set of ordinary differential equations in a mean field picture as described in Intra-host HIV model with latency.

We are thus considering latent and actively infected cells that become active with probability  . Birth rates are given by
. Birth rates are given by 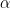 s and deaths by
s and deaths by  s. Activation is given by
s. Activation is given by 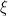 and the remaining variables are typical but expressed using greek letters.
and the remaining variables are typical but expressed using greek letters.
Note, we can be clear on where the viruses all go if we use  then, using the quasi equilibrium
then, using the quasi equilibrium 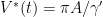 . If the latent pool is disregarded, the basic reproduction number
. If the latent pool is disregarded, the basic reproduction number  is calculated when the population is fully susceptible, i.e.
is calculated when the population is fully susceptible, i.e.  , so we solve the equation
, so we solve the equation

which gives the solution, defining 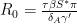
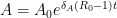
which goes to zero precisely when 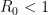 . The result then is when the basic reproduction number is less than one, the virus will always die out with probability equal to one.
. The result then is when the basic reproduction number is less than one, the virus will always die out with probability equal to one.
In the situation where the virus does die out, calculating the generations until the die out is useful. We formulate the branching process stochastic analogue to the ODEs above considering only activated and latent cells. We assume the original number of active cells is then  . The active cell can then make virions which go on to infect other cells. Thus we have the possible transitions:
. The active cell can then make virions which go on to infect other cells. Thus we have the possible transitions:

assuming that the number of susceptible cells is at equilibrium .
The probability of 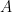 cells arising from
cells arising from 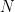 viruses assuming a single virus infects a single cell is then
viruses assuming a single virus infects a single cell is then
 .
.
Probability generating functions
With the probability, we can then write the probability generating function (pgf, as described in my post: Adding beans to a pot) for the probability of active cells as

putting in the probability  , we can group the terms with exponent , and use the binomial theorem
, we can group the terms with exponent , and use the binomial theorem  . This leads to
. This leads to

so that defining  we have
we have
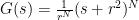
We might take to also be a random variable because active cells can produce varying numbers of virions, . The probability distribution  arises from the sequence of events that leads to virions before the cell dies. Thus the probability distribution is the product of birth events having normalized probability
arises from the sequence of events that leads to virions before the cell dies. Thus the probability distribution is the product of birth events having normalized probability  and a single event with probability
and a single event with probability  :
:

We can then take the average generating function with respect to the probability of , in the discrete sense, this is a sum over all multiplied by so that we have
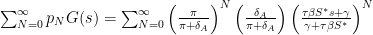
so we can group terms with exponent , and then use that the sum of the geometric series  for $latex|x| \leq 1$. This leads to the average generating function with respect to , i.e.
for $latex|x| \leq 1$. This leads to the average generating function with respect to , i.e.  :
:
![\langle G(s) \rangle_N = \left(\frac{\delta_A}{\pi+\delta_A}\right) \left[1-\left( \frac{\tau\beta S^*s + \gamma}{\gamma + \tau\beta S^*} \frac{\pi}{\pi+\delta_A} \right)\right]^{-1}](https://s0.wp.com/latex.php?latex=%5Clangle+G%28s%29+%5Crangle_N+%3D+%5Cleft%28%5Cfrac%7B%5Cdelta_A%7D%7B%5Cpi%2B%5Cdelta_A%7D%5Cright%29+%5Cleft%5B1-%5Cleft%28+%5Cfrac%7B%5Ctau%5Cbeta+S%5E%2As+%2B+%5Cgamma%7D%7B%5Cgamma+%2B+%5Ctau%5Cbeta+S%5E%2A%7D+%5Cfrac%7B%5Cpi%7D%7B%5Cpi%2B%5Cdelta_A%7D+%5Cright%29%5Cright%5D%5E%7B-1%7D&bg=%23ffffff&fg=%23000000&s=0&c=20201002)
or pulling the terms into the denominator
![\langle G(s) \rangle_N = \left[\frac{\pi+\delta_A}{\delta_A}-\frac{\tau\beta S^*s + \gamma}{\gamma + \tau\beta S^*} \left(\frac{\pi}{\delta_A} \right)\right]^{-1}](https://s0.wp.com/latex.php?latex=%5Clangle+G%28s%29+%5Crangle_N+%3D+%5Cleft%5B%5Cfrac%7B%5Cpi%2B%5Cdelta_A%7D%7B%5Cdelta_A%7D-%5Cfrac%7B%5Ctau%5Cbeta+S%5E%2As+%2B+%5Cgamma%7D%7B%5Cgamma+%2B+%5Ctau%5Cbeta+S%5E%2A%7D+%5Cleft%28%5Cfrac%7B%5Cpi%7D%7B%5Cdelta_A%7D+%5Cright%29%5Cright%5D%5E%7B-1%7D&bg=%23ffffff&fg=%23000000&s=0&c=20201002)
Now factoring

we can cancel the first and last terms in the denominator

factoring again we have

Conway defines  so that the average generating function is finally
so that the average generating function is finally

Extinction process
We now have the expression for the generating function

If we know an extinction will happen, that is, is less than 1, we have a subcritical branching process. We can consider the probability of an infected cell producing  offspring as
offspring as  , where we assume a maximum amount does exist.
, where we assume a maximum amount does exist.
The probability of ultimate death (extinction) in the  th generation we call
th generation we call  . For
. For  , or extinction in the th generation, requires everything to die after the
, or extinction in the th generation, requires everything to die after the  th generation. The probability of
th generation. The probability of 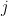 cells dying in generation
cells dying in generation  is
is  . For all birth events to die out then we must balance each birth probability with the equivalent death probability, i.e.:
. For all birth events to die out then we must balance each birth probability with the equivalent death probability, i.e.:

for which the rhs is precisely the pgf we defined before. That means we have the condition

Now let’s try to solve this relationship for the definitions of  given above, identifying
given above, identifying  . Conway gives us
. Conway gives us 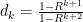 , where I’ve dropped the subscript. Let’s check that with the average pgf:
, where I’ve dropped the subscript. Let’s check that with the average pgf:
First 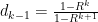

cross multiplying

totally writing all terms out
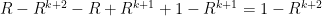
we can see immediately that the equality is true. Therefore, we have derived the probability of dying out in the k-th generation.
Note this result only works for  otherwise we could have negative probabilities. In this “subcritical” regime, we can be interpreted the expression as the probability of not extinction in the next generation relative to probability of not-extinction in some generation.
otherwise we could have negative probabilities. In this “subcritical” regime, we can be interpreted the expression as the probability of not extinction in the next generation relative to probability of not-extinction in some generation.

