
The canonical model
I end up doing calculations related to this model quite a bit, so I’m leaving it here for myself and others.
I’ll begin with what I refer to as the “canonical model of HIV dynamics”. This is an ordinary differential equation set that governs susceptible cells 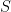
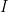

So, the equation set is typically written
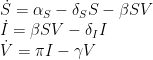
in which the over-dot notation indicates time derivatives (e.g. 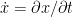
So, we have the constant birth rate 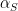


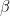


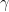
A lot of HIV cure research these days is focused on removing “latent” HIV. This is a state where HIV has integrated its DNA into a cell, but the cell is not producing virus, thus the immune system (or medicine, more on that in a bit) does not see the virus, and doesn’t remove it. We are going to model the latent state too.
We expand the infected compartment of the canonical model to account for the latently infected cells 
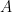

We also separate out the dynamics because evidence shows latently infected cells live like uninfected cells, dividing to make daughter cells and dying naturally—also note there will be cool dynamics because these cells are in fact CD4+ T cells which are immune system cells in themselves and likely clonally expand to fight other pathogens… but we ignore that for now. We let latent cells proliferate and die with rates 


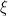
At this point too we include HIV medicine. Antiretroviral therapy as it is called is incredibly effective at stopping viral replication to the point that modern ART allows HIV-infected patients to have undetectable viral levels, and live a normal life. Still, motivation for “cure” persists because if a patient cannot take the medicine strictly, like for example if they live in a place where it is hard to get, a latent cell can reactivate and restart the infection. In the model, we denote the effectiveness of ART therapy ![\epsilon \in [0,1]](https://s0.wp.com/latex.php?latex=%5Cepsilon+%5Cin+%5B0%2C1%5D&bg=%23ffffff&fg=%23000000&s=0&c=20201002)

Our new set of equations is then
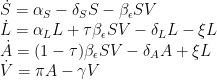
Equilibrium solutions of the model
Equilibrium solutions to the set of ODEs above can be calculated by setting all the time derivatives to zero. We denote the equilibrium state with an asterisk).
First we solve the fourth equation for 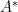
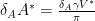
then identify 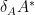

then solving the second equation for 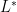

so that we can factor and cancel

![\frac{\delta_A \gamma}{\pi} = \left[\frac{-\xi\tau}{\theta_L}+(1-\tau)\right]\beta_\epsilon S^*](https://s0.wp.com/latex.php?latex=%5Cfrac%7B%5Cdelta_A+%5Cgamma%7D%7B%5Cpi%7D+%3D+%5Cleft%5B%5Cfrac%7B-%5Cxi%5Ctau%7D%7B%5Ctheta_L%7D%2B%281-%5Ctau%29%5Cright%5D%5Cbeta_%5Cepsilon+S%5E%2A&bg=%23ffffff&fg=%23000000&s=0&c=20201002)
and rewrite the bracketed term as 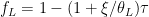
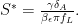
From here, we solve for the equilibrium viral load concentration, or the viral set-point using the first equation

and the others follow, leading to the set of equilibrium solutions:
![S^* = \frac{\gamma\delta_A}{\beta_\epsilon \pi f_L} \\ L^* = \frac{\tau}{\theta_L}\left[\frac{\gamma\delta_S\delta_A}{\beta_\epsilon \pi f_L}-\alpha_S\right]\\ A^* = \frac{\alpha_S f_L}{\delta_A} - \frac{\gamma\delta_S}{\beta_\epsilon \pi}\\ V^* = \frac{\alpha_S\pi f_L}{\gamma\delta_A}- \frac{\delta_S}{\beta_\epsilon}](https://s0.wp.com/latex.php?latex=S%5E%2A+%3D+%5Cfrac%7B%5Cgamma%5Cdelta_A%7D%7B%5Cbeta_%5Cepsilon+%5Cpi+f_L%7D+%5C%5C+L%5E%2A+%3D+%5Cfrac%7B%5Ctau%7D%7B%5Ctheta_L%7D%5Cleft%5B%5Cfrac%7B%5Cgamma%5Cdelta_S%5Cdelta_A%7D%7B%5Cbeta_%5Cepsilon+%5Cpi+f_L%7D-%5Calpha_S%5Cright%5D%5C%5C+A%5E%2A+%3D+%5Cfrac%7B%5Calpha_S+f_L%7D%7B%5Cdelta_A%7D+-+%5Cfrac%7B%5Cgamma%5Cdelta_S%7D%7B%5Cbeta_%5Cepsilon+%5Cpi%7D%5C%5C+V%5E%2A+%3D+%5Cfrac%7B%5Calpha_S%5Cpi+f_L%7D%7B%5Cgamma%5Cdelta_A%7D-+%5Cfrac%7B%5Cdelta_S%7D%7B%5Cbeta_%5Cepsilon%7D&bg=%23ffffff&fg=%23000000&s=0&c=20201002)

1 Comment